酶
具有高度专一性的催化活性的蛋白质。1878年以前称为酵素,以后改称为酶。1926年首次将脲酶制备成结晶,并证明它是蛋白质。以后又研究了上百种酶,证明酶都是蛋白质。
生物体内的代谢反应,几乎都是由酶催化的。由于酶是生物体内的催化剂,所以又称酶为生物催化剂。受酶催化进行化学变化的物质称底物。由酶催化的化学反应称酶促反应。迄今已知的酶已达2000种以上。有些酶只有一条肽链,但多数酶由两条以上的肽链组成,每条肽链称为它的一个亚基。有些酶是单纯蛋白质,有些酶则是结合蛋白质。结合蛋白质酶的分子中,除蛋白质外,还含有非蛋白质的辅助因子。辅助因子可以是金属离子(如Fe2+、Mg2+、Mn2+等),也可以是小分子有机物,这类酶只有当酶蛋白与其辅助因子结合成为全酶时才具有酶活性。那些和酶蛋白松弛结合的有机辅助因子称为辅酶;结合紧密,不易分离者称为辅基。辅酶和辅基可作为电子或某种化学基团的供体或受体而参与酶促反应。许多维生素是构成辅酶和辅基的成分。
酶的命名与分类 按照国际系统命名原则,每种酶有一个系统名称和习惯名称。习惯命名通常以酶的来源、底物或催化的反应机制为依据。例如来源于胃的水解蛋白质的酶称胃蛋白酶。习惯名称简单, 使用方便, 但容易混淆。系统名称则是根据酶的底物名称及催化的反应类型而命名。例如乳酸脱氢酶是习惯名称, 其系统名称是乳酸NAD氧化还原酶。系统名称较长, 但不会混淆。依据国际系统分类法, 将酶分为6大类:氧化还原酶类; 移换酶类; 水解酶类; 裂合酶类; 异构酶类和合成酶类。6大类分别用1,2,3,4,5,6的编号来表示。再根据底物中被作用的基团或键的特点, 将每一大类分为若干个亚类, 亚类亦按顺序编成1,2,3,4……等。每个亚类再分为若干个次亚类,仍用1,2,3,4……编号。次亚类中每个酶也按顺序编号。因此每个酶都有一个由4个数字组成的分类编号, 数字间由“·”隔开。第一个数字表示该酶属于6大类中哪一类; 第二个数字表示该酶属于哪一亚类; 第三个数字表示该酶属于哪一个次亚类; 第四个数字则表示该酶在次亚类中的排列号。在编号前面往往还冠以酶学委员会的缩写E.C.(Enzyme Commission)。例如乳酸:NAD氧化还原酶的分类编号是E.C.1.1.1.27。
酶的作用机制 在化学反应中, 反应物的分子必须具有足够的能量, 即达到或超过该反应所特需的能量时, 才能成为活化态(过渡态)并发生反应。活化态比初始态高出的能量称为活化能。反应物各个分子的能量不同, 有的处于活化态, 能够发生反应, 有的则否。所以化学反应的速度取决于反应物活化分子在分子总数中所占的比例。酶能降低底物分子的活化能,因而使活化分子的比例大大增加,从而加快了反应速度。酶降低活化能的原因是在酶促反应中酶(E)先和底物(S)结合成不稳定的中间产物(ES), 然后中间产物(ES)迅速裂解成稳定的产物(P), 并游离出酶。其反应可用下式表示
E+S⇌ES→E+P
由于中间产物ES的形成,使原来的反应途径(S→P) 发生了改变, 而新反应途径的活化能比较低, 所以反应速度加快。由于酶的分子大而底物的分子小,所以仅由酶分子表面的局部区域与底物结合, 此区域称酶的活性部位或活性中心。酶的活性部位由结合部位和催化部位所组成。结合部位决定与何种底物结合,催化部位决定催化反应的性质。单纯蛋白质组成的酶活性部位是由肽链中某些氨基酸残基形成的三维结构。由结合蛋白质组成的酶的活性部位除氨基酸残基外,还含有辅基的某些基团。酶活性部位有严格的空间构象, 这是由其一级结构决定的。任何改变活性部位空间构象的因素都影响酶的活性。
酶作用的专一性 酶和其他催化剂不同, 对它所作用的底物有严格的选择性,一种酶只能催化某一类,甚至某一种物质发生一定的化学变化, 称为酶作用的专一性或特异性。酶的专一性分为3类: ①有些酶只催化一种底物发生一定变化, 而不作用于任何其他物质,称为酶的绝对专一性。例如脲酶只催化尿素水解,而对尿素的各种衍生物(如尿素的甲基取代物)都不起作用。②有些酶的专一性略低, 它能催化一类底物发生反应, 这种专一性称为酶的相对专一性。这类酶只对底物的某一化学键起作用, 而对键两端基团的要求程度不同, 有的仅对其中一个基团要求严格, 有的则对键两端的基团都无严格要求。③立体构型专一性。这类酶只对立体异构型(旋光异构和顺反异构)中的某一构型起作用,而对另一构型则不起作用。例如L-氨基酸氧化酶只催化L-氨基酸氧化, 而对D-氨基酸无作用。1890年德国化学家费歇尔(E. Fisher)提出“钥匙—锁”假说来解释酶作用的高度专一性。他认为底物分子的一部分必须像钥匙那样, 能够专一地契入与它互补的酶表面(锁), 才能产生催化作用, 因而一种酶只能催化具有特定形状的分子发生反应, 所以酶有高度专一性。然而后来发现有些酶的活性部位是可塑的,当底物分子与酶结合时, 在底物分子的诱导下, 使酶活性部位的形状改变为与底物分子互补而结合, 此即诱导契合学说。当酶从中间复合物(ES)中解离出来后,酶即恢复原来的形状。
影响酶促反应速度的因素 酶促反应速度受多种因素影响。在有足够底物的情况下,酶促反应速度(V)与酶(E)浓度成正比, V=K〔E〕, K为反应速度常数。当酶浓度、温度和pH恒定时, 在底物浓度很低的范围内, 反应的初速度几乎与底物浓度成正比, 当底物浓度达一定程度后, 所有酶都结合上了底物, 反应速度达到最大值(Vm), 此时, 再增加底物浓度也不能使反应速度增加。1913年德国生物化学家米埃利斯和门腾(L. Michaelis & M. Menten) 根据中间产物的概念, 以方程式定量的表示了酶促反应速度与底物浓度的关系: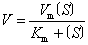
式中 Vm即反应最大速度; Km为米氏常数;〔S〕为底物浓度。此公式与许多酶的实际情况相符, 从而成为中间产物学说的有力证明。在达到最大反应速度一半时,底物浓度等于Km。大多数酶的Km值在10-7~10-1范围内。酶能最有效地发挥作用的温度称酶的最适温度。动物和人体内酶的最适温度一般在35~40℃之间。植物酶的最适温度稍高, 在40~50℃之间。高温能使酶变性, 故使酶的活力降低, 甚至失活。低温时酶稳定,其活力也低。所以在实践中,常在低温下保存酶制剂以及某些含酶的生物材料。
酸碱度影响酶或底物的解离,从而影响酶的活性。在最适pH条件下,酶促反应的速度最大, 高于或低于此pH值,反应速度均下降。酶的激活剂能提高酶活力。
酶的抑制剂使酶的活力下降。抑制剂和激活剂与酶结合后,会改变酶的构象,从而影响酶的活力,但并不引起酶蛋白变性。酶的抑制剂有多种。有的结构与底物相似,它们和底物竞争与酶的活性部位结合,因而阻碍底物与酶结合,减少酶的作用机会,使酶活力降低。有的抑制剂和酶结合,并不影响酶与底物结合,但酶的催化活性受到抑制。在上述两种抑制作用中,抑制剂与酶蛋白都是以非共价键结合,所以可用透析法除去抑制剂,使酶恢复活性,称为可逆性抑制作用。磺胺类药物(如对氨基苯磺胺)的结构和对氨基苯甲酸十分相似,后者是细菌中催化二氢叶酸合成的二氢叶酸合成酶的底物。由于磺胺药物是此酶的竞争性抑制剂,它抑制了二氢叶酸的合成,因而使细菌的繁殖受到抑制。有些抑制剂能与酶的活性部位的某些基团共价结合,使酶失去活力。由于这类抑制剂不能用透析、超滤等物理方法除去,因而称这种作用为不可逆抑制作用。例如有机磷化合物(农药1059、敌百虫等)不可逆的抑制与中枢神经活动有关的胆碱酯酶,使乙酰胆碱不能分解为乙酸和胆碱,乙酰胆碱在组织内积存,引起神经中毒症状。有机汞和有机砷化合物也是由于不可逆的抑制了生物体内含硫氢基的酶而产生中毒作用。
酶活性测定 测定酶活性是在特定的最适条件下(最适pH、最适温度、适当的底物浓度和排除抑制剂等)测定酶促反应体系中单位时间内产物的生成量或底物的消耗量,并以反应的速度表示酶的活性。应尽可能测定反应开始时的速度(初速),以免除种种干扰。酶活性的高低用国际单位表示,一个酶活性单位是指在酶作用的最适条件下(最适pH、最适底物浓度)25℃,一分钟催化1.0微克分子底物发生转化的酶量。比活性是指每毫克蛋白质所具有的酶活性。比活力愈高,表明酶愈纯。检测血清或组织样品中某些酶活性的方法,常应用于诊断疾病。正常情况下,某些酶由细胞释放到血液中的量很少,血清中含量较低。患病时,有些酶从受损伤的细胞中释放到血液中,使血清中的含量增加,酶活性升高。例如肝炎和心肌梗塞时,血清中谷—丙转氨酶活性常明显增加;猪患应急综合症,血清中磷酸肌酸激酶活性升高;患骨质疏松症时,血清中碱性磷酸酶含量升高等。
酶原 本身不表现催化活性,经过激活后可转变为酶的一类蛋白质。例如胰脏分泌的胰蛋白酶原及胰凝乳蛋白酶原在小肠内活化成胰蛋白酶和胰凝乳蛋白酶。活化过程是酶原分子被切断一个或多个肽键,使其构象发生一定的变化,从而成为具有催化活性的酶。这种活化过程通常是在酶的催化下进行的。如肠激酶将胰蛋白酶原转变为胰蛋白酶。活化的酶也可催化同种酶原的活化,如胃蛋白酶可以激活胃蛋白酶原等。细胞最初合成的是酶原,然后在适当条件下激活为酶是有重要意义的。例如许多在消化道中起作用的蛋白酶在细胞内合成的是无活性的酶原,这样不致分解细胞内的蛋白质。而分泌到消化道内以后,被激活而发挥消化作用。又如正常存在于血液中的凝血酶原不起凝血作用,一旦血管破裂被激活为凝血酶,便产生促进凝血的作用。

